Fatty Acid Oxidation (FAO)
FAO has been implicated in the control of eating since the first demonstration of an increase in food intake in response to intraperitoneal injection of the FAO inhibitor mercaptoacetate (MA). For more than 20 years the prevailing view was that MA stimulates eating by inhibiting hepatic FAO, thus activating a vagal afferent signal from the liver. More recently this concept has been questioned because (1) hepatic parenchymal vagal afferent innervation is scarce and because (2) experimentally induced changes in hepatic FAO often fail to produce changes in eating. Therefore, one goal of our research in this context is to test the novel alternative hypothesis that changes in small intestinal epithelial cell (=enterocyte) FAO influence eating. More specifically, we investigate (1) whether fatty acids derived from ingested dietary fat can be oxidized in enterocytes, (2) whether this process generates a chemical mediator that is released from enterocytes and (3) affects vagal afferent firing, thus providing a signal to the brain that influences eating. The working hypothesis is depicted in Figure 1 (Langhans et al., Am J Physiol 300:R554-R565, 2011). We address these questions by employing sophisticated experimental surgical, behavioral, transgenic, electrophysiological and analytical techniques in a combination of in-vivo studies in spontaneously eating rats and mice as well as in-vitro studies in cell cultures and tissue preparations.
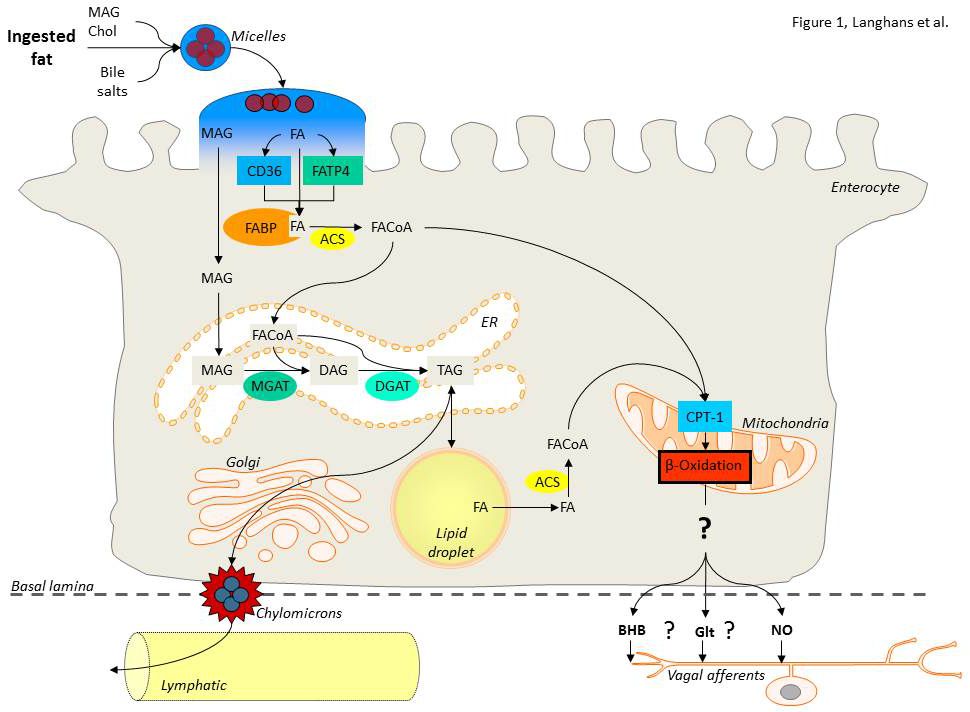
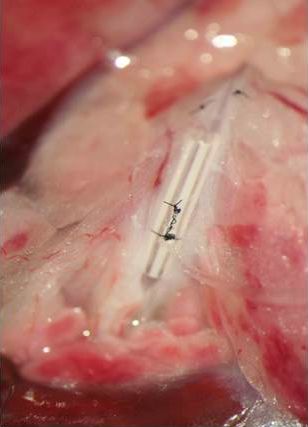
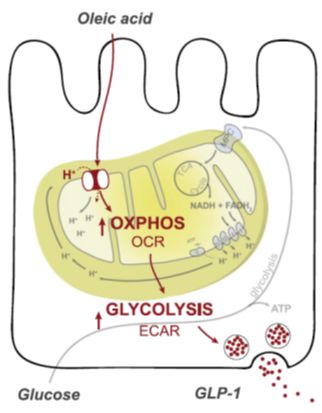
Figure 2 shows another unique technique developed in our laboratory, i.e., the cannulation of the intestinal lymph duct in rats. This technique allows for the repetitive collection of intestinal lymph in freely behaving animals, which is particularly useful for investigations into the mechanisms of enterocyte fat handling. Also, we also address the role of fatty acid oxidation in the stimulation of GLP-1 release (see Figure 3 for a summary of the pertinent experimental data; Clara et al., Metabolism 65:8-17, 2016).
The basic research significance of this research is that it will provide novel information about peripheral metabolic mechanisms that influence eating, an issue that is still largely unresolved. Given the multiple side effects and other problems frequently encountered with anti-obesity drugs acting in the brain, and promoted by the dramatic obesity-curbing effects of bariatric surgery, attention in obesity research is slowly shifting from the brain to the periphery, in particular to the gastrointestinal tract and to the gut-brain communication. In providing new information about a potentially powerful intestinal metabolic mechanism of eating control, this research has substantial translational significance. Most recently, we started to expand the investigations addressing the role of FAO in the control of eating to include a possible role of astrocytes, which are often considered to be hepatocytes in the brain.